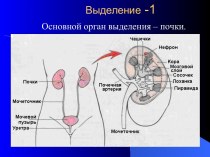
")






























![МОЗЖЕЧОК.новая часть [кора наружной зоны полушарий + зубчатые ядра] – автоматизация произвольных](/img/tmb/15/1405155/8f4b1999c6868c13675b914fa01a00e4-720x.jpg "Двигательные рефлексы спинного мозга. Локомоция: центры конечностей, тонические и фазические управляющие влияния")











особому расписанию:
Группы 1-2 – с 9-00 до 9-55 Группы
3-4 – с 10-00 до 10-55Группы 5-6 – с 11-00 до 11-55 Группы 7-8 – с 12-00 до 12-55
Кроме тестов по лекциям 11-14, каждый желающий 16 мая сможет переписать (или написать – при пропуске по неуважительной причине) еще 2 теста по лекциям 1-10.
Пропустившие семинары по уважительной причине (наличие справ-ки) могут, кроме того, написать соответствующие тесты (в этом случае они должны прийти к своему преподавателю в группы 1-4).
Пропустившие более 2-х тем (более одного семинара) по неуважительной причине должны будут в день экзамена сдать лектору письменные (рукописные) работы по пропущенным и «неотработанным» 16 мая темам (ответы на 40 вопросов, распо-ложенных в конце каждой лекции). Допуск к дальнейшему экзамену – при условии успешной сдачи письменных работ лектору.